I funghi
🍄 I funghi sono fonte di sostentamento per noi e per tutto ciò da cui dipendiamo. Scopriamoli insieme!


I funghi sono ovunque, ma è facile non notarli. Sono dentro e fuori di noi. Sono fonte di sostentamento per noi e per tutto ciò da cui dipendiamo.
Merlin Sheldrake
Caratteristiche
I funghi (dal latino “fungus”), chiamati anche miceti (dal greco “mykes”), costituiscono un regno di organismi molto dissimili tra loro, ma che hanno in comune le seguenti proprietà:
- Come gli animali, le pianti e i protisti, sono eucarioti, ossia la loro unità fondamentale è la cellula eucariotica, ovvero dotata di un nucleo che racchiude il materiale genetico (DNA);
- Attraverso specifici enzimi, si nutrono di vari substrati organici presenti in natura (eterotrofi);
- Si adattano ad ambienti con ossigeno (aerobi obbligati o facoltativi) o privi di esso (anaerobi);
- Possono essere formati da una sola cellula o da più cellule (unicellulari o pluricellulari).
Nell’immaginario comune i miceti sono riconosciuti come “funghi eduli e non”, lieviti e “muffe”. I primi ricordano maggiormente i vegetali, sono infatti i tipici funghi con gambo e cappello (carpoforo) che possiamo trovare quando passeggiamo nei boschi. Ovviamente sappiamo tutti che alcuni di essi sono velenosi, come Amanita phalloides (tignosa verdognola), Amanita muscaria (ovolo malefico), Boletus satanas (porcino malefico) e Russola emetica (colombina rossa), mentre altri sono mangerecci, come Boletus edulis (porcino), Cantharella cibarius (gallinaccio), Macrolepiota procera (mazza di tamburo), Agaricus bisporus (champignon) e Armillaria mellea (chiodino). A proposito dei funghi del genere Armillaria, forse non tutti sanno che l’organismo più grande del mondo è il micete Armillaria ostoyae, che si estende nel sottosuolo per circa 965 ettari (9.650.000 mq). Un fungo da record!
I lieviti, ben noti per il loro utilizzo nell’ambito enologico o nella produzione di birre, sono costituiti da una singola cellula di forma sferoidale o ellissoidale (unicellulari). Si possono vedere soltanto al microscopio e, come altri funghi, se fatti crescere nei terreni di coltura, creano delle colonie visibili a occhio nudo. Il più conosciuto è Saccharomyces cerevisiae, noto come “lievito di birra”, che è solitamente impiegato per la preparazione di vino, birra, pane e pizza.
Le “muffe” sono i miceti che si sviluppano su frutta, verdura, pane, vegetali, muri e tappezzerie. Chiamate anche ifomiceti o funghi filamentosi, esse si presentano come degli agglomerati di strutture filamentose, tubulari e pluricellulari (dette ife) che generano degli ammassi visibili al livello macroscopico. Le ife sono contraddistinte da ramificazioni e intrecciamenti, e insieme formano il micelio. Tutti i funghi che presentano un micelio e che producono spore senza il carpoforo sono definiti “muffe” nel parlare comune. Per fare qualche esempio, citiamo Neurospora crassa (muffa del pane), Cladosporium cladosporioides (cresce su carta, pasta di legno, vernici, affreschi, fibre tessili e davanzali bagnati), Aspergillus niger (muffa nera su frutta e verdura) e Penicillium roqueforti (muffa commestibile dei formaggi come il gorgonzola).
Tuttavia, questa classificazione porta a delle incongruenze quando si entra nel dettaglio a parlare di specifici funghi o di alcuni loro stadi. Per esempio, i cosiddetti “funghi eduli” non sono altro che una struttura prodotta da alcuni miceti per riprodursi, mentre il “vero” fungo è un’intricata rete di ife (che nella classificazione comune sarebbe definita “muffa”). Nel paragrafo sulla tassonomia verrà spiegata la classificazione utilizzata in ambito scientifico e che aiuta a comprendere e suddividere meglio i miceti.
I funghi sono creature complesse che si possono trovare in tutti gli ambienti. Infatti, risiedono in svariati luoghi come l’acqua, il suolo, le foglie, i frutti, i detriti vegetali, i tronchi degli alberi, ma anche negli animali. Essendo così ubiquitari, si adeguano ai diversi ambienti con facilità e si specializzano. Solitamente preferiscono un’umidità elevata (90%-100%) e un pH basso (6.0-6.8). Chiaramente possono esistere eccezioni che fanno si che i funghi riescano a prosperare in condizioni fuori dal comune, come nei deserti o in terreni con un ampio intervallo di pH, dato che sono organismi che si adattano a condizioni notevolmente diverse e, a volte, estreme.
Per quanto concerne la temperatura, distinguiamo tre categorie di miceti: psicrofili (crescono a temperature inferiori a 0° C, con una temperatura ottimale di 15° C, e sopportano un massimo di 20° C), termofili (si sviluppano da una temperatura di 20° C fino ad un massimo di 62 °C) e mesofili. Questi ultimi hanno un intervallo di temperatura di crescita che si accavalla con gli psicrofili e i termofili, cioè dai 5° C ai 10° C fino ad un massimo di 35-40° C; ciò che cambia è la temperatura migliore per riprodursi.
I funghi, data l’assenza di clorofilla (se non in condizioni di simbiosi mutualistiche), sono incapaci di effettuare la fotosintesi. Per tale ragione la loro crescita non necessita della luce solare, anzi alcuni funghi prediligono l’assenza o una bassa quantità di luce, e ciò gli permette di proliferare in molti più ambienti possibili. Di conseguenza, i miceti hanno bisogno di substrati per nutrirsi, e si alimentano mediante assorbimento.
Le cellule fungine
Le principali tipologie di cellule fungine sono le ife (le più diffuse), le cellule di lievito e le spore. In generale la cellula fungina è composta dalla parete cellulare, la membrana, il citoscheletro, il vacuolo, il nucleo, il citoplasma e i vari organelli (Fig. 1).
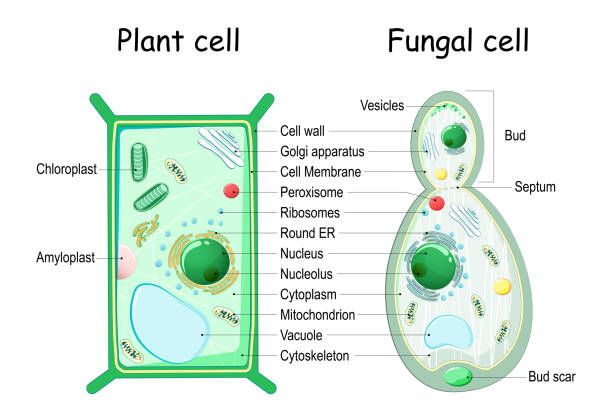
Iniziamo dalla parete cellulare, che raffigura il collegamento dinamico tra l’organismo e l’ambiente. Oltre a questo, ha diversi compiti, i più importanti sono: determinare e mantenere la morfologia del fungo, l'integrità delle cellule, permettere alla cellula fungina di generare un alto turgore, proteggere le cellule dai cambiamenti della pressione osmotica esterna e da altri stress ambientali. La parete appare come una struttura multistratificata costituita da un reticolo compatto di zuccheri complessi o polisaccaridi (chitina, glucani, mannani e galattomannani), lipidi, proteine e glicoproteine (proteine unite agli zuccheri). Essendo i funghi creature molto complesse, è da specificare che la composizione precisa, tuttavia, varia notevolmente tra le diverse specie di miceti, è altamente regolata e sensibile ai cambiamenti ambientali. Grazie alla membrana plasmatica al di sotto della parete, avviene uno scambio regolato di elementi e sostanze chimiche. La membrana è formata da un doppio strato di fosfolipidi (una tipologia di lipidi complessi) in cui sono inseriti steroidi e glicoproteine.
Il citoscheletro è costituito da tre elementi: microtubuli, microfilamenti di actina e filamenti intermedi, che si estendono nel citoplasma, dal nucleo alla membrana plasmatica. Questa struttura ha molteplici funzioni, ossia permette alle cellule di cambiare la loro forma, rinforzare la membrana plasmatica e quella nucleare, trasporta le vescicole nel citoplasma (grazie a delle proteine motrici), costituisce il fuso mitotico (essenziale per la divisione cellulare) e consente il movimento di alcuni organelli.
Passiamo ora alla descrizione dello spazio interno della cellula, che è riempito dal citoplasma, una sostanza acquosa contenente zuccheri, amminoacidi, proteine, acidi grassi, sali minerali, e in cui sono immersi i vari organuli, ognuno circondato da una membrana. L’organulo principale è il nucleo, dentro al quale è collocato il DNA, che forma i cromosomi; a seconda della cellula che si considera, può essere singolo o multiplo.
Gli altri organuli sono i mitocondri (essenziali per il ciclo dell’acido citrico, l’ossidazione dei grassi e la produzione di molecole energetiche), il reticolo endoplasmatico liscio (dove avviene la genesi dei lipidi), il reticolo endoplasmatico rugoso (dove vengono sintetizzate le proteine), l’apparato di Golgi (che gioca un ruolo importante nella maturazione delle proteine, nella sintesi dei glicolipidi di membrana e nell’assemblaggio dei polisaccaridi della parete) e i vacuoli. Questi ultimi hanno molteplici ruoli: la regolazione dell’omeostasi cellulare, il traffico di membrana, la segnalazione, la nutrizione, lo stoccaggio di polifosfati e amminoacidi, la rimozione di composti tossici dal citoplasma e l'autofagia (processo attraverso cui le cellule degradano i propri organuli). Nel caso dei funghi, questi organelli acidi sono simili sia a quelli delle piante che ai lisosomi dei mammiferi, e hanno una morfologia che varia a seconda del tipo di micete, dalla cellula e dall’età.
Come detto all’inizio, i principali tipi di “cellule” sono:
- Ife, che è scorretto definirle cellule. Esse sono costituite da strutture complesse e filamentose, e hanno combinazioni uniche di attributi strutturali, comportamentali e funzionali. Per esempio, l’ifa vegetativa solitamente è multinucleata e presenta dei setti (delle pareti trasversali); grazie alla sua crescita apicale e la produzione di diramazioni, permette l’allargamento della colonia fungina e la sintesi del micelio (creando una complessa e interconnessa rete ifale);
- Cellule di lievito, che sono tipicamente cellule singole. Si riproducono attraverso un processo caratterizzato dalla creazione di un setto che porta alla divisione della cellula madre e alla conseguente produzione di due cellule figlie. Questo avviene sia per gemmazione (Saccharomyces cerevisiae) che per fissione binaria (Schizosaccharomyces pombe). Alcuni patogeni sono definiti lieviti dimorfi perché in condizioni particolari possono generare delle strutture allungate chiamate pseudoife (come Candida albicans). Inoltre, alcuni lieviti, come Cryptococcus neoformans, formano una capsula di polisaccaridi attorno a una parete cellulare melanizzata. Questa capsula conferisce ai lieviti la possibilità di diventare “cellule titano” aumentando di una ventina di volte il loro diametro;
- Spore, che sono strutture cellulari discrete prodotte da quasi tutti i funghi. La loro forma varia a seconda del fungo che le procrea e possono essere unicellulari o, come risultato della settatura, multicellulari. Non a caso, i tipi di spore e le strutture che rilasciano spore, tra cui i corpi fruttiferi pluri-ifali, sono utilizzate per la classificazione e l’identificazione dei funghi.
Quindi in generale, per semplificare la complessità di questo organismo, si può dire che i funghi sono caratterizzati da un corpo, il tallo, che può essere unicellulare o pluricellulare, e la loro organizzazione cellulare è di tipo eucariota, ovvero sono composti da cellule che comprendono specifici componenti, dei quali il più rimarchevole è il nucleo.
Tassonomia e riproduzione
La riproduzione dei miceti è molto complessa e viene utilizzata al fine di classificarli a livello tassonomico; per tale ragione la procreazione e la tassonomia verranno spiegate insieme. Solitamente la riproduzione si fonda sulla sporulazione, ovvero la genesi e l’emissione nell’ambiente di una tipologia di cellule dette spore, le quali possono originarsi per via sessuale o asessuale, che si intervallano durante il ciclo vitale fungino; nel primo caso parliamo di meiospore (come le ascospore, le zigospore, le basidiospore), nel secondo di mitospore (ad esempio le sporangiospore, i conidi e le clamidospore). I funghi che presentano entrambe le tipologie di riproduzione (asessuale e sessuale) sono detti telomorfi o perfetti, mentre quelli di cui non è nota attualmente la riproduzione sessuale sono detti anamorfi o imperfetti. C’è da sottolineare che l’impiego delle spore da parte dei funghi ha la stessa funzione dell’utilizzo dei semi da parte delle piante: lo scopo è quello di diffondersi.
Prima di esporre i due meccanismi di riproduzione, è necessario fornire una descrizione riguardante la sporulazione, che si verifica in tre passaggi:
- Liberazione, che consente alle spore di staccarsi dal fungo che le ha generate, e ciò può avvenire in maniera attiva o passiva. La prima implica che le cellule al di sotto della spora perdano acqua e si raggrinziscano, l’acqua viene sostituita dall’aria inducendo il rigonfiamento e il conseguente scatto nella cellula che proietta le spore. La seconda, invece, è favorita dalla gravità, dalle correnti dell’aria o dall’acqua;
- Dissipazione, durante la quale le spore vengono trascinate dalle correnti e arrivano a percorrere distanze anche grandi. Ci sono molteplici fattori che influenzano la disseminazione, per esempio la forma, la dimensione, la rugosità, la carica elettrica delle spore, la temperatura, la viscosità dell’aria, l’umidità, le correnti aeree e il moto del vento;
- Adagiamento, cioè il momento in cui la spora raggiunge uno specifico substrato, si deposita e può andare incontro a vari destini: maturare e in seguito germinare, oppure entrare in uno stato di quiescenza (dormienza, torpore). La maturazione consiste in una serie di cambiamenti morfologici che rendono la spora in grado di germinare; durante questa fase la spora assorbe le sostanze nutritive presenti sul substrato (carboidrati, grassi, amminoacidi), aumenta di volume, produce chitina, cellulosa e proteine, e inizia a formare il tallo. La quiescenza è una condizione reversibile in cui le spore rimangono inattive e non proseguono il loro ciclo, e tale stato è causato da fattori ambientali come la carenza di umidità, le temperature troppo alte o troppo basse e la mancanza di nutrienti. Quando le condizioni ambientali diventano più favorevole, la spora può uscire dalla dormienza e svilupparsi.
Le forme di riproduzione differenziano i sei phyla (o divisioni) tassonomici dei funghi:
- Chytridiomycota: i funghi appartenenti a questo phylum si distinguono per la presenza di zoospore con una singola struttura posteriore simile a una frusta (il flagello), che permette la mobilità. La maggior parte di questi miceti si trova in acqua dolce o in terreni umidi e, prevalentemente, sono parassiti di alghe, piante (Chytridiales spp.) e animali (Batrachochytrium dendrobatidis negli anfibi) ed altri funghi;
- Blastocladiomycota: come i Chytridiomycota, sono funghi acquatici che producono zoospore flagellate (poco comune nei funghi). Ciò che distingue questo phylum dagli altri è che le spore hanno un unico flagello e possiede un singolo grande nucleo diploide circondato da un “cappuccio nucleare” composto da un assemblaggio di ribosomi legati tra loro. Risiedono nell’acqua dolce, nel fango e nel suolo, e sono saprofiti e patogeni delle piante (Physoderma deformans e Physoderma maydis, che infettano rispettivamente l’anemone e il mais);
- Neocallimastigomycota: sono funghi anerobi obbligati che solitamente si trovano nel rumine o nel cieco degli animali erbivori, ed aiutano gli ospiti nella digestione della cellulosa. Si riproducono per via asessuata e, anche loro, producono zoospore mobili con uno o molti flagelli. I talli sono monocentrici o policentrici. Queste spore sono caratterizzate dall’assenza di mitocondri, ma idrogenosomi (che usano il piruvato e il malato per produrre idrogeno e anidride carbonica);
- Glomeromycota: sono dei miceti strettamente legati al suolo che creano associazioni micorriziche con le piante, e solitamente fanno parte delle endomicorrize (all’interno dell’ospite). Sono dei simbionti obbligati che producono spore di grandi dimensioni (40-800 μm), multistrato, attaccate a delle ife non settate. Alcuni esempi sono Glomus spp., Acaulospora spp. e Sclerocystis spp.;
- Ascomycota: è il gruppo più grande e più vario di funghi esistenti. La caratteristica principale di questo phylum è l’asco, una struttura a sacco dove vengono prodotte le ascospore. Però non tutti i funghi appartenenti a questa divisione, seppure in forma minore anche alle altre, si riproducono sessualmente. Per questo motivo sono suddivisi in funghi perfetti (cioè che producono gli aschi) e funghi imperfetti (o deuteromiceti) che hanno una riproduzione asessuale. La classificazione di quest’ultime avviene per analisi genetiche e tra queste ci sono tre classi principali: Saccharomycotina (usualmente lieviti che si riproducono per gemmazione), Taphrinomycotina (sono di norma dei simil-lieviti e patogeni delle piante) e Pezizomycotina (comprende la maggior parte dei funghi imperfetti ascomiceti).
Alcuni esempi di funghi appartenenti a questo phylum sono: Penicillium spp., Aspergillus spp., Trichoderma spp., Neurospora spp., Morchella spp. (nota come spugnola), Gyromitra esculenta (falsa spugnola), Saccharomyces spp., Candidaspp., i tartufi e numerosi funghi patogeni degli animali e delle piante;
- Basidiomycota: è il gruppo dei classici funghi che vediamo nei boschi e alcuni dei miceti più dannosi per le piante. Questo phylum è caratterizzato dalla produzione di spore sessuali su una struttura chiamata basidio (simile ad una clava), che spesso è collocata sopra o all’interno di un corpo fruttifero chiamato basidiocarpo. La riproduzione asessuata avviene con la gemmazione diretta delle basidiospore oppure delle ife con la produzione di conidi. In questo gruppo possiamo riconoscere i funghi commestibili e i miceti tossici riportati all’inizio, e in più sono inclusi anche Fomes fomentarius, Heterobasidium annosum, Trametes versicolor, Ganoderma (lucidum e applanatum), Dichomitus squalens, Serpula lacrimans, Gloephyllum trabeum e Grifola frondosa.
C’è da sottolineare che la tassonomia dei funghi è tutt’oggi non ben definita, tant’è che alcuni dei phyla sopra citati vengono considerati dei sub-phyla, e altri ricercatori aggiungono alla lista anche gli Zoopagomycota, che sono miceti parassiti di amebe, insetti (Entomophtora muscae e Zoophagus pectosporus) e piccoli animali, ma anche di altri funghi, e i Mucoromycota, che sono possono essere sia funghi strettamente legati al suolo, ma anche dannosi per l’essere umano o le piante; due esempi sono Rhizopus stolonifer (muffa nera del pane) e Absidia spp. (agente patogeno delle mucormicosi). In altri casi, i phyla Zoopagomycota e Mucoromycota sono considerati sub-phylum degli Zygomycota.
È importante far presente che la tassonomia è in continuo cambiamento, per esempio recentemente dai phylum sono stati rimossi i seguenti organismi, che però vengono ancora gestiti come se fossero funghi:
- Microsporidia: definiti più simil-funghi che miceti veri e propri, sono caratterizzati da spore minute, che contengono un materiale infettivo conosciuto come sporoplasma. Comunemente si trovano nell’intestino degli insetti (Nosema bombycis nei bachi da seta) e nei muscoli dei pesci. In linea di massima sono considerati patogeni, infatti, quando le spore vengono ingerite da un nuovo ospite, gli organismi si incistano nell'epitelio intestinale e raggiungono tessuti specifici attraverso il sangue. Raggiunte le cellule interessate, iniziano a dividersi ripetutamente in maniera asessuata. Alla fine, raggiunto lo stadio maturo (trofozoiti), danno origine a zigoti prodotti sessualmente che genereranno nuove spore;
- Oomyceti: appartengono alla classificazione superiore degli Heterokonta e non dei fungi, ma per lungo tempo sono stati trattati come miceti, dato che hanno caratteristiche simili. Creano sporangi contenenti zoospore biflagellate (un flagello posteriore ed uno anteriore). La diffusione delle spore avviene con film d’acqua o in acqua. Di questa classificazione fa parte la Phytophtora infestans (nota come la peronospora delle patate).
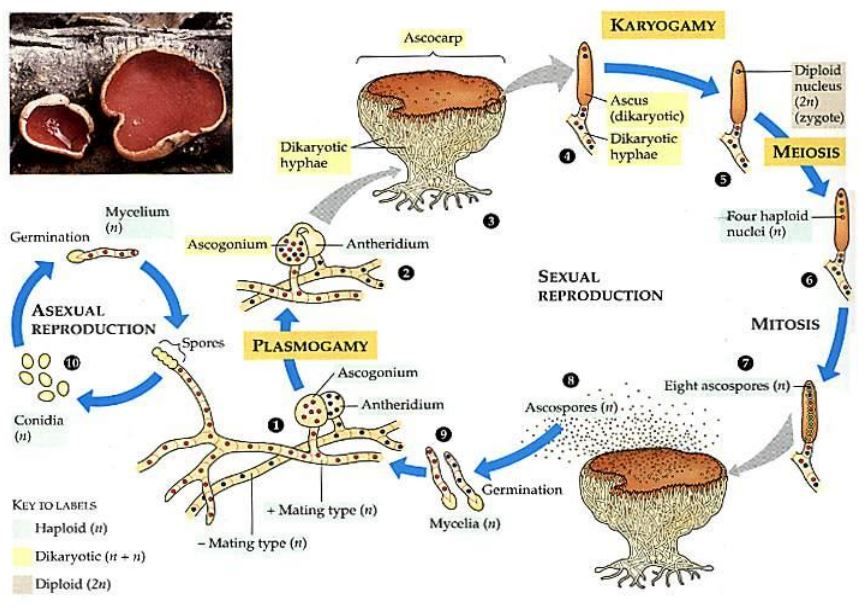
La riproduzione fungina è molto complessa e si suddivide in riproduzione sessuata e asessuata (Fig. 2 e 3). La prima avviene mediante la creazione di spore diploidi (con 48 cromosomi nel nucleo) chiamate conidi. Si tratta di mitospore che si formano in seguito a uno specifico processo di divisione cellulare (mitosi) da parte dei conidiofori, che sono localizzati all’estremità delle ife fungine. L’emissione dei conidi può essere seguita da due processi dissimili, ovvero gemmazione (tipica dei lieviti) e frammentazione delle ife (caratteristica delle muffe); il primo consiste nella scissione di una cellula primaria (cellula madre) che porta alla formazione di cellule più piccole (cellule figlie o blastospore), le quali possono staccarsi dalla madre oppure rimanere legate ad essa. Può capitare che le blastospore si dividano in maniera successiva e restino unite tra loro generando le pseudoife. Nel secondo processo il conidio si differenzia in filamenti che si accrescono, si ramificano e si intrecciano fino a giungere alla realizzazione del micelio, che è organizzato in porzione aerea (che si trova a contatto con l’aria e svolge una funzione riproduttiva rilasciando spore) e porzione vegetativa (che rimane a contatto col terreno e assimila i nutrienti). Questa disposizione del micelio si osserva non solo nelle muffe, bensì anche nei funghi eduli, dove il cappello e il gambo non sono altro che i prodotti della fruttificazione del micelio localizzato nel terriccio e della compattazione dei filamenti del micelio aereo; la parte inferiore del cappello è deputata al rilascio delle spore.
Passiamo alla riproduzione sessuata, che consiste nella ricombinazione del materiale genetico di due cellule, al fine di creare organismi geneticamente dissimili da quelli originari. Questa tipologia di riproduzione normalmente si verifica in condizione avverse e si può differenziare in due tipi di accoppiamento: omotallico (autofertile) oppure eterotallico (tra due miceli diversi, ma compatibili). In generale, la riproduzione sessuale si può distribuire in tre fasi sequenziali: plasmogamia, cariogamia e meiosi. Le ife producono i cromosomi diploidi che vengono separati in due cellule figlie aploidi (24 cromosomi). Successivamente avviene la plasmogamia dove le due cellule aploidi si fondono, diventando una cellula dicarionte con entrambe i nuclei aploidi in un’unica cellula. Durante la cariogamia i due nuclei si uniscono e generano uno zigote diploide, il quale si divide per meiosi formando nuclei aploidi che vengono incorporati in spore chiamate meiospore; queste, dopo la maturazione, si diffondono. Una volta trovato il substrato adeguato, in condizioni vantaggiose di temperatura e umidità, esse iniziano a germinare garantendo lo sviluppo del fungo. Data la complessità dei funghi, il processo della plasmogamia può essere differente:
- Cellule sessuali specializzate (gameti) che vengono rilasciate da organi sessuali differenziati chiamati gametangi;
- Due gametangi entrano in contatto e i nuclei passano dal gametangio maschile a quello femminile, assumendo così la funzione di gameti. Questo si realizza soprattutto negli ascomiceti, dove il gametangio maschile e femminile sono detti anteridio e ascogonio. Le spore dicariotiche sono raccolte a formare un asco dal quale verranno poi disperse le ascospore;
- Fusione dei gametangi per riunire i loro nuclei;
- Produzione di ife somatiche che assumono la funzione sessuale ed entrano in contatto, si fondono e si scambiano i nuclei. Solitamente si compie nei basidiomiceti. Le ife dicariotiche (che raffigurano il micelio secondario) si addensano e creano il micelio terziario dal quale si forma il basidiocarpo o carpoforo (il “classico” fungo dei boschi). Quest’ultimo porterà i basidi, di norma nel cosiddetto “cappello”, che rilasceranno le basidiospore.

Ecologia
I funghi dipendono dalla materia organica presente in natura, quindi, sono organismi eterotrofi. Oltre all’enorme complessità di questo regno, tale proprietà rende ancora più difficile studiare la loro ecologia. Si può generalizzare dicendo che, allo scopo di acquisire gli elementi nutritivi, i miceti adottano quattro strategie diverse: saprofitismo, commensalismo, mutualismo e parassitismo.
Il saprofitismo consiste nel cibarsi di materiale organico in putrefazione, per esempio i tronchi degli alberi secchi, le foglie, gli escrementi, le carcasse e l’humus. I funghi saprofiti hanno un ruolo ecologico importantissimo: degradando la sostanza organica morta, cioè scomponendola in sali minerali, restituiscono nutrimenti al suolo. Se ciò non avvenisse, la grossa mole di foglie e residui vegetali che si depositano sul terriccio sommergerebbero gli alberi fino a soffocarli. Tra le altre cose, è stato osservato che maggiore è la biodiversità nel terreno, più sarà efficiente la produzione di humus per nutrire il suolo.
I miceti possono essere considerati dei “super decompositori”; infatti, grazie ad un gruppo di specifici enzimi, sono in grado di scomporre sostanze organiche solitamente difficili da “digerire”, come il legno. Questi funghi hanno una dieta varia e sono costituiti da diverse popolazioni, quindi, prenderemo come esempio uno dei numerosi substrati che possono decomporre, cioè la lettiera del bosco, e riporteremo alcuni miceti che svolgono tale compito. Le principali sostanze bersaglio sono l’amido, la cellulosa, la pectina e la lignocellulosa.
L’amido è il polisaccaride usato come riserva energetica nelle piante. Il suo deterioramento porta all’ottenimento di zuccheri semplici come il maltosio e il glucosio.
La cellulosa e la pectina sono due polisaccaridi che rappresentano componenti della parete delle cellule vegetali. Alla fine della degradazione della cellulosa, si avranno degli zuccheri, mentre, nel caso della pectina, degli acidi e degli alcoli.
La lignocellulosa è una combinazione di tre zuccheri complessi (cellulosa, emicellulosa e lignina). Questo polimero è il legno, ossia l’“impalcatura” delle piante. Solo in Europa si possono contare circa 2700 funghi che si nutrono di questo substrato. Il legno, sia vivo che morto, può subire tre tipologie di degradazione causate solitamente da diversi funghi basidiomiceti, che sono responsabili di tre patologie:
- La carie bianca che colpisce sia conifere che latifoglie. I funghi responsabili distruggono la lignina ed arrivano alla cellulosa e all’emicellulosa, che si trovano immerse nella matrice della lignina. Il legno, in seguito alla degradazione, assume delle tonalità più chiare e una consistenza fibrosa. Alcuni funghi di questo gruppo sono Fomes fomentarius, Trametes versicolor, Dichomitus squalens e Pleurotus spp;
- La carie bruna è causata da funghi che degradano la cellulosa e l’emicellulosa, ma non la lignina (seppure la lascino solo parzialmente intatta), dal momento che non sintetizzano un determinato enzima: l’esoglucanasi. Ciò rende il legno colpito più friabile, meno resistente e con una colorazione scura. A tale fazione appartengono Serpula lacrimans, Gloephyllum trabeum e Fomitopsis pinicola (poliporo marginato). Il carpoforo di quest’ultimo può superare i 40 cm di larghezza e i 20 cm di spessore, oltre ad essere un gigante dei boschi; non a caso è considerato uno dei più forti degradatori delle foreste di conifere;
- La carie soffice, che consiste nell’alterazione contemporanea e lenta di lignina e cellulosa, generando un ammorbidimento dei tessuti per permettere l’attacco di altri organismi saprofiti. Alcuni esempi sono Alternaria spp. e Phialophora spp., che indeboliscono diverse piante e non solo il legno.
Il commensalismo è una forma di simbiosi in cui due organismi si associano e vivono in stretto contatto; uno ricava nutrienti essenziali dall’altro organismo senza però arrecargli alcun danno o sofferenza. L’esempio migliore di commensalismo è l’insieme dei microrganismi (funghi e batteri) che fanno parte del microbiota umano, ovvero che vivono in diverse parti del nostro organismo (bocca, naso, faringe, intestino, cute, vie urinarie, organi sessuali). Tra i funghi commensali umani abbiamo i lieviti Malassezia furfur, Trichosporon cutaneum e Candida spp.; i primi due risiedono solo sulla cute, mentre il terzo si trova sulla cute, nel tratto gastroenterico, genitourinario e respiratorio. In alcune circostanze (per esempio in presenza di immunodepressione), questi funghi possono indurre l'insorgenza di patologie che verranno trattate nei prossimi paragrafi
Il mutualismo è un’altra tipologia di simbiosi in cui entrambi gli organismi associati traggono beneficio, come nel caso dei licheni e delle micorrize.
I licheni sono il risultato di una relazione tra un fungo (organismo eterotrofo definito micobionte) e un’alga verde e, in alcuni casi, anche un cianobatterio (organismo autotrofo che sarà il fotobionte). Tale struttura è una semplificazione della complessità di questi organismi, dove si possono osservare simbiosi con batteri, funghi e alghe. Nel corpo che si crea il micobionte è collocato nella zona più esterna, mentre il fotobionte si trova nella parte più interna. Tra i due avviene uno scambio reciproco di sostanze, ovvero il fungo procura all’alga o al cianobatterio l’acqua e i sali minerali che assorbe dal substrato, mentre il fotobionte fornisce al fungo i carboidrati prodotti mediante la fotosintesi. L’unione dei due genera un organismo che al livello morfologico e strutturale non somiglia a nessuno dei due componenti; questo perché le sue proprietà non derivano dalla semplice fusione delle caratteristiche del micobionte e di quelle del fotobionte, bensì sono l’esito di una co-evoluzione che ha permesso la genesi di un organismo capace di vivere in maniera indipendente, laddove il fungo o l’alga da soli non riuscirebbero. Facciamo l’esempio degli acidi lichenici, che vengono sintetizzati solamente dal lichene, e non dal fungo o dall’alga allo stato libero. Questa simbiosi permette ai licheni di colonizzare molteplici substrati che spaziano da organici ad inorganici (il suolo, gli alberi, i muschi, l’asfalto, il cuoio, il cemento, i mattoni, le tegole e il vetro), e per fare ciò adattano anche la loro morfologia: i licheni fruticosi presentano un corpo ramificato che somiglia a un cespuglio, i licheni crostosi hanno l’aspetto di croste e sono attaccati saldamente al substrato, infine i licheni fogliosi sono distinti da lobi appiattiti che ricordano delle foglie e crescono parallelamente al substrato. Per fare un paio di esempi, nelle passeggiate in montagna potreste aver visto la Cladonia spp. (lichene fruticoso) su degli alberi abbattuti o nei suoli ricchi di humus, oppure il Pleopsidium chlorophanum (lichene crostoso) su rocce o pareti rocciose.
Le micorrize si formano dall’associazione tra alcuni funghi del terreno e le radici non lignificate di alcune piante. Anche in questo caso c’è un organismo eterotrofo ed uno autotrofo, e tra di essi si verifica uno scambio di nutrienti: le ife fungine, che sono molto più sottili e lunghe rispetto alle radici vegetali, si conficcano nel suolo più in profondità, scavano nelle rocce attraverso la produzione di acidi, ricavano i sali minerali (specialmente azoto e fosforo) e li trasferiscono alla pianta con cui sono associati. Questa funzione, per esempio, è vitale per le piante nel deserto. La pianta, tramite la fotosintesi, sintetizza gli zuccheri che fungono da nutrimento per il micete. Nella loro connessione, le piante e i funghi micorrizici raffigurano polarità contrastanti, in quanto i germogli delle piante interagiscono con la luce e l’aria, mentre i funghi e le radici con il terriccio. Dobbiamo rimarcare che i funghi micorrizici sono altamente fruttuosi, a tal punto che il loro micelio costituisce tra un terzo e la metà degli organismi viventi che popolano il terreno. I contatti tra ife e radici non sono statici, bensì si rimodellano in continuazione, e inoltre i funghi micorrizici creano delle ampie reti miceliali che mettono in connessione le piante (Wood Wide Web) consentendo lo scambio di materiali tra di esse. Per fare un esempio di micorrize, in particolare ectomicorrize (all’esterno dell’ospite), non possiamo non citare i tartufi, che sono i corpi fruttiferi prodotti sottoterra di una specifica tipologia di funghi micorrizici, come il tartufo bianco (Tuber magnatum) e il tartufo nero (Tuber melanosporum).
Il parassitismo indica una forma di interazione in cui un organismo (parassita) si colloca sopra o all’interno di un altro organismo (ospite) e vive a sue spese; di conseguenza in questo tipo di relazione il parassita trae beneficio, mentre l’ospite soffre e viene danneggiato. I funghi parassiti sono quelli associati a malattie negli animali (patogeni) e nelle piante (fitopatogeni). Alcuni miceti patogeni sono parassiti obbligati, altri a seconda delle condizioni esterne possono anche essere parassiti o saprofiti. I funghi patogeni dell’essere umano si possono suddividere in quattro gruppi in base alla tipologia di infezioni (micosi) che causano, ovvero superficiali, cutanee, sottocutanee e profonde.
Le micosi superficiali coinvolgono gli strati più esterni dell’epidermide, i peli e i capelli. Due funghi di questo gruppo sono gli ascomiceti Malassezia furfur, responsabile della tinea versicolor, i cui segni clinici sono la comparsa di macchie cutanee rosa o marrone chiaro (dovute alla ipopigmentazione o iperpigmentazione), e Piedrai hortae, un fungo saprofita del suolo che provoca la piedra nera, distinta da noduli scuri e compatti su peli o capelli.
Le micosi cutanee (dermatomicosi) attaccano le zone del corpo ricche di cheratina come l’epidermide, le unghie, i peli e i capelli. I responsabili sono i cosiddetti funghi dermatofiti o cheratinolitici (cioè che degradano la cheratina). In questo gruppo sono inclusi tre generi di muffe parassite obbligate appartenenti al phylum Ascomycota, cioè Microsporumspp., che privilegia la cute e i capelli, Trichophyton spp., che invade la cute, le unghie e i capelli, ed Epidermophytum spp., che contagia la cute e le unghie. Questi miceti provocano il quadro clinico definito dermatofitosi o tinea, che si localizza in diverse parti del corpo, tra cui lo scalpo (tinea capitis), le zone della barba (tinea barbae), l’inguine (tinea cruris), le mani (tinea mannum), i piedi (tinea pedis), i siti dove la pelle è liscia e glabra (tinea corporis), e le unghie (tinea unguim od onicomicosi). Le lesioni cutanee si presentano simili a dei brufoli che poi si allargano in maniera centrifuga e formano dei cerchi rossastri lasciando intatta la cute al centro. Le unghie infette appaiono scolorite, rialzate, fragili e deformate. A livello dei capelli e dei peli, questi funghi si sviluppano sulla superficie esterna (Microsporum) o interna (Trichophyton):nel primo caso si parla di infezione ectotrica, che induce una caduta reversibile del pelo o del capello, mentre nel secondo caso l’infezione è endotrica e la perdita pelo o del capello è irreversibile.
Ci sono delle micosi cutanee che non sono provocate dai miceti dermatofiti, bensì dal lievito commensale Candida, che in alcune situazioni promuove l’insorgenza di infezioni (candidosi) che possono essere superficiali o profonde. In questa parte ci concentriamo su quelle superficiali, di cui abbiamo la candidosi intertriginosa, la dermatite da pannolino, la candidosi ungueale, la candidosi vulvovaginale e il mughetto. Le prime due colpiscono la cute e si manifestano con arrossamento, desquamazione e prurito, la terza interessa le unghie, che sono distinte da lisi, screpolatura, annerimento e rilievo dei bordi, la quarta interessa la mucosa vaginale, è caratterizzata da arrossamento, prurito e secrezioni di colore bianco, e l’ultima si concentra nella bocca, dove si formano delle placche biancastre.
Le micosi sottocutanee e profonde sono causate specialmente da funghi che hanno la caratteristica di crescere sotto forma di muffe a temperatura ambiente (25° C), mentre si trasformano in lieviti a temperature di 37° C (dimorfi). Quando si trovano nell’ambiente esterno, sono saprofiti e assumono la forma miceliare che rilascia spore, ma nel momento in cui le spore entrano all’interno di un organismo ospite (mediante inalazione o traumi cutanei), acquisiscono l’aspetto lievitiforme, vivono come parassiti e innescano patologie. Per esempio, l’ascomicete Sporothrix schenckii, che risiede soprattutto nelle piante, e di solito penetra in un organismo ospite in seguito a punture, graffi o tagli quando si manipolano piante contaminate. La malattia (sporotricosi) colpisce i tessuti sottocutanei e i linfonodi, ed è caratterizzata dalla formazione di papule o noduli.
Le micosi profonde insorgono in seguito all’inalazione delle spore, inizialmente colpiscono i polmoni ma dopo possono diffondersi ad altri organi determinando malattie sistemiche che possono risultare fatali. Alcuni funghi responsabili sono gli ascomiceti Blastomyces dermatitidis, Histoplasma capsulatum, Coccidioides immitis e Paracoccidioides brasiliensis. Questi in natura si collocano nel terreno, nelle piante, nel legno in putrefazione e negli escrementi di animali, dove vivono come saprofiti.
Per quanto concerne i funghi fitopatogeni, determinano la comparsa di un vasto numero di patologie nelle piante, che danneggiano sia la qualità dei prodotti che la resa della produzione, e potenzialmente arrecano danno secondario all’essere umano (schianto degli alberi). Prendiamo come esempi tre ascomiceti e due basidiomiceti:
- Venturia inaequalis (Fig. 4A), agente della ticchiolatura del melo, che si manifesta con macchie pigmentate sulle foglie, sui frutti e sui fiori;
- Sclerotinia sclerotiorum (Fig. 4B), che è l’agente eziologico della muffa bianca su una vasta gamma di piante, tra cui soia, girasole, colza, pomodoro, leguminose, cucurbitacee e ombrellifere. Questo micete crea un micelio bianco e vellutato e determina ingiallimento degli organi verdi, avvizzimento e caduta delle foglie;
- Botrytis cinerea (Fig. 4C), responsabile della muffa grigia su kiwi, fragole e vite. I fiori infetti si scuriscono e si seccano, con conseguente blocco della riproduzione della pianta, mentre se il micete colpisce i frutti, questi diventano molli e compare la massa filamentosa grigia.
- Heterobasidium annosum (Fig. 5A), che causa i marciumi radicali e la carie bianca delle radici e della porzione basale del tronco di alberi di conifere (abete, larice, pino silvestre e pino cembro). L’infezione delle radici permette al micete di propagarsi in altri alberi con conseguente diffusione a macchia d’olio della patologia. Gli alberi infetti presentano scolorimento della corteccia, marcescenza delle radici e dei gambi ed eccessiva produzione di resina;
- Fomes fomentarius (Fig. 5B), un patogeno di alcune latifoglie come faggio, pioppo e acero, su cui induce la marcescenza del legno o carie bianca. L’infezione comporta il marciume, la colorazione chiara del legno, la comparsa di linee scure e cavità colme di micelio, fino alla morte dell’albero infetto. Dopo la morte, il micete continua a vivere sul tronco e da parassita diventa saprofita.


Funghi medicinali
Questa sezione la dedichiamo ai miceti che possiedono proprietà medicinali, ovvero che contengono sostanze che apportano beneficio alla nostra salute. Per tale ragione vengono impiegati nella medicina naturale che, in questo caso, prende il nome di micoterapia. I funghi che descriveremo sono tre basidiomiceti saprofiti, cioè Hericium erinaceus, Ganoderma lucidum e Trametes versicolor (Fig. 6):
- Hericium erinaceus (detto anche testa di scimmia o criniera di leone) è diffuso nelle aree montane dell’emisfero boreale e prolifera su alberi di latifoglie morti come la quercia, il faggio e il noce. Questo fungo è commestibile ed esercita un’azione protettiva sul sistema nervoso grazie alla presenza di molecole steroidee chiamate erinacine, le quali stimolano la produzione del fattore neurotrofico, una proteina che promuove la crescita e la sopravvivenza dei neuroni, favorisce i processi rigenerativi, riduce il danno ischemico cerebrale e inibisce l’infiammazione. Il suo utilizzo potrebbe prevenire malattie neurodegenerative come il morbo di Alzheimer e il morbo di Parkinson, e si è rivelato utile efficace negli individui affetti da sclerosi multipla. In aggiunta a ciò, H. erinaceus, grazie a degli zuccheri (beta-glucani) potenzia il sistema immunitario, con effetto protettivo verso alcune infezioni batteriche (per esempio da Salmonella ed Helicobater pylori). In più abbassa l’infiammazione a livello gastrointestinale, con conseguente miglioramento nelle persone affette da colite ulcerosa o morbo di Chron;
- Ganoderma lucidum (Reishi in Giappone), conosciuto come “fungo dell’immortalità”, è distribuito in tutto il mondo e si sviluppa, specialmente nei periodi più afosi, su alberi di latifoglie come castagno, quercia, faggio, olmo e acero. Include una cospicua mole di sostanze benefiche, tra cui gli acidi ganoderici, molecole steroidee che limitano il rilascio di istamina (una sostanza prodotta nelle reazioni allergiche) e abbassano i livelli di colesterolo LDL e trigliceridi. Inoltre, contiene i beta-glucani e una proteina specifica (LZ-8), che hanno attività modulante sul sistema immunitario, consentendo una maggiore difesa verso le infezioni e i tumori. G. lucidum di per sé non è commestibile, infatti viene trattato al fine di ottenere l’estratto e/o la polvere e impiegarlo come integratore alimentare. In commercio lo si può trovare sotto forma di capsule o in polvere, oppure inserito all’interno di prodotti alimentari come cioccolato, caffè e tè;
- Trametes versicolor (denominato anche Turkey Tail, coda di tacchino o poliporo a mensola variopinto) è un fungo non mangereccio da cui si estrae uno zucchero legato a una proteina, chiamato polisaccaride K, che è contenuto nel farmaco Krestin, impiegato come coadiuvante nella chemioterapia, dal momento che blocca le cellule neoplastiche e stimola il sistema immunitario. L’uso di questo micete nella micoterapia abbraccia la prevenzione e la terapia dei tumori ormono-dipendenti, l’epatite, l’herpes, le infezioni, le malattie respiratorie, il supporto del sistema immunitario, il trattamento degli effetti collaterali della chemioterapia e la radioterapia, la stanchezza, le malattie croniche e le fibromialgie. Come G. lucidum, anche T. versicolor viene venduto sotto forma di integratore alimentare in capsule.

Funghi e alimentazione
Vediamo ora alcuni aspetti dell'utilizzo dei funghi in ambito culinario senza, tuttavia, soffermarci troppo su quelli scontati e che quasi tutti conoscono (es. porcini, chiodini, champignon etc.). Piuttosto descriveremo dei funghi più particolari e meno comuni.
Perché mangiarsi un piatto di chiodini o di porcini quando possiamo mangiarci un huitlacoche (Fig. 7A)? Stiamo parlando di Ustilago maydis, un basidiomicete parassita che danneggia i chicchi di mais, provocando la malattia chiamata carbone del mais, caratterizzata da ingrigimento e anomalo rigonfiamento dei chicchi. Potrebbe sembrare stravagante, ma in Messico le pannocchie contaminate dal fungo (dette huitlacoche) raffigurano un ingrediente di molteplici ricette come il succotash (un piatto a base di mais dolce e fagioli), la quesadilla (tortilla di mais farcita), le empanadas (fagottini di pasta ripieni di verdure o carne), i taco (tortilla di farina di mais o di frumento, ripiegata e farcita in vari modi), le frittate e i risotti. C’è da dire che le huitlacoche sono conosciute dai tempi antichi, dal momento che erano incluse nella cucina azteca. Sono distinte da un sapore terroso, sono ricche di sotolone (zucchero lattone), vanillina e glucosio, e contengono una notevole concentrazione di proteine, più alta rispetto alle pannocchie non infette.
Un altro micete peculiare è Pleurotus ostreatus (chiamato anche fungo orecchione o fungo ostrica) (Fig. 7B), un basidiomicete saprofita che cresce su alberi di latifoglie, soprattutto pioppi e gelsi. Si tratta di un micete dalla carne compatta e tenera, dal sapore dolciastro e l’odore gradevole. Può essere coltivato facilmente in due modi, ovvero mettendo in una fossa uno strato di micelio insieme a dei tronchi di pioppo tagliati (coltivazione “manuale” a terra), oppure usando un substrato già preparato, acquistabile da casa, formato da paglia, mais, grano e piante leguminose, su cui seminare le colture di micelio (coltivazione “industriale”). Considerando l’ottima commestibilità e la coltivazione semplice, P. ostreatus gode di un vantaggio gastronomico rilevante. Pertanto può essere cucinato in multiple maniere, ovvero gratinato, impanato, trifolato, in padella, sott’olio, farcito, essiccato e polverizzato, bollito e condito con olio. Oltre alle proprietà culinarie, P. ostreatusha anche delle qualità terapeutiche, dal momento che contiene la lovastatina, una sostanza in grado di abbassare i livelli di colesterolo, e gli zuccheri beta-glucani.
Sappiamo che un bicchiere di vino ad ogni pasto fa bene, e qui ritorna il lievito Saccharomyces cerevisiae, che ha una proprietà che viene sfruttata in ambito alimentare, ossia il micete trae nutrimento dagli zuccheri, li metabolizza e rilascia specifiche sostanze di scarto: in presenza di ossigeno produce anidride carbonica (CO₂), e ciò favorisce la crescita degli impasti per pane e pizza (lievitazione), poiché la CO₂ rimane intrappolata nell’impasto. In assenza di ossigeno, invece, converte gli zuccheri in etanolo (fermentazione alcolica), consentendo quindi la produzione di vino, birra e sidro.
Concludiamo parlando dei funghi come fonte di proteine, un’alternativa più sostenibile rispetto alla carne animale. Introduciamo quindi il concetto di “micoproteine”, che indica non solo la parte proteica, bensì l’intero corpo del micete; il fungo maggiormente utilizzato per la creazione di micoproteine su scala industriale è Fusarium venenatum (Fig. 7C), un ascomicete filamentoso che si trova nel suolo. Le proteine ricavate da questo fungo sono già in commercio con il marchio Quorum. Le micoproteine hanno delle buone proprietà nutrizionali, dal momento che, oltre alle proteine, contengono fibre (che sono dei prebiotici per la flora intestinale), vitamine del gruppo B e acidi grassi saturi e polinsaturi, il cui rapporto è pari a 1:3. In aggiunta a ciò, permettono un controllo dei picchi di glucosio e insulina dopo i pasti.

Lorenzo Tramontana
Fonti
G. Pasqua, G. Abbate e C. Forni. 2019. “Botanica generale e diversità vegetale (IV edizione)”, Piccin
Merlin Sheldrake. 2020. “L’ordine nascosto. La vita segreta dei funghi”, Marsilio Editori
Kathleen K. Treseder, Renaud Berlemont , Steven D. Allison, Adam C. Martiny. 2018. “Drought increases the frequencies of fungal functional genes related to carbon and nitrose acquisition”, PLOS One
Johannes Rousk, Erland Bååth, Philip C Brookes, Christian L Lauber, Catherine Lozupone, J Gregory Caporaso, Rob Knight & Noah Fierer. 2010. “Soil bacterial and fungal communities across a pH gradient in an arable soil”, International Society for Microbial Ecology
M. Wang, X. Jiang, W. Wu, Y. Hao, Y. Su, L. Cai, M. Xiang and X. Liu. 2015. “Psychrophilic fungi from the world’s roof”, Persoonia
Saroj Ahirwar, Hemant Soni, Bhanu Pratap Prajapati & Naveen Kango. 2017. “Isolation and screening of thermophilic and thermotolerant fungi for production of hemicellulases from heated environments”, An International Journal on Fungal Biology
Zhi-Yan Du, Krzysztof Zienkiewicz, Natalie Vande Pol, Nathaniel E Ostrom, Christoph Benning and Gregory M Bonito. 2019. “Algal-fungal symbiosis leads to photosynthetic mycelium”, eLife
Tatiana Yu. Gagkaeva, Aleksandra S. Orina, Olga P. Gavrilova, and Nadezhda N. Gogina. 2020. “Evidence of microdochium fungi associated with cereal grains in Russia”, Microorganisms
2019. “Scientists identify two new species of fungi in retreating Arctic glacier”, ScienceDaily https://www.sciencedaily.com/releases/2019/01/190115121103.htm
Nicholas P. Money. 2016. “Fungal diversity”, The Fungi (terza edizione)
Thomas N. Taylor. 2009. “Fungi, Bacteria and Lichens”, The biology and evolution of fossil plants, paleobotany (seconda edizione)
Jules J. Berman. 2012. “Taxonomic guide to infectious diseases. Chapter 36 - Ascomycota”
Thomas J Volk. 2001. “Fungi”, Encyclopedia of Biodiversity
Heikki Setälä e Mary Ann McLean. 2004. “Decomposition rate of organic substrates in relation to the species diversity of soil saprophytic fungi”, Oecologia
Fry William E. and Niklaus J. Grünwald. 2010. “Introduction to Oomycetes”, The Plant Health Instructor
Baggio J.S., Goncalves F.P., Lourenco S.A., Tanaka A.O., Pascholati S.F., Amorim L. 2016. "Direct penetration of Rhizopus stolonifer into stone fruits causing rhizopus rot", Plant Pathology
Jenna Battillo. 2018. “The role of corn fungus in Basketmaker II diet: A paleo nutrition perspective on early corn farming adaptations” Science Direct
Martina Murdolo. 2012. “Medicina di laboratorio”, Universitalia
Yashvant Patel, Ram Naraian and V.K. Singh. 2012. “Medicinal properties of Pleurotusspecies (Oyster mushroom): A review”, World Journal of Fungal and Plant Biology
https://www.istitutoagrariosartor.edu.it/wp-content/uploads/2013/11/I-funghi.pdf
“I funghi”, mondofunghi.com https://www.mondofunghi.com/
“Il ruolo essenziale per legno morto e vecchio”, vogelwarte.ch https://www.vogelwarte.ch/it/atlas/focus/il-ruolo-essenziale-per-legno-morto-e-vecchio#:~:text=Il%20legno%20morto%20%C3%A8%20di,alpestre%20o%20Cincia%20dei%20salici
“Ecologia dei funghi”, dolomiti.it https://www.dolomiti.it/it/funghi/ecologia-dei-funghi
https://terapistioccupazpisa.files.wordpress.com/2011/01/micologiase09.pdf
"Lignocellulosa: l'innovazione per i biocarburanti", agronotizie.imagelinetwork.com https://agronotizie.imagelinenetwork.com/bio-energie-rinnovabili/2009/02/10/lignocellulosa-l-innovazione-per-i-biocarburanti/32092
“La necromassa: la seconda vita del faggio”, forestbeat.it http://www.forestbeat.it/la-necromassa/
“Ascomiceti”, mondofunghi.it https://www.mondofunghi.com/ascomiceti
http://shigen.nig.ac.jp/algae_tree/NeocallimastigomycotaE.html
“Glomeromycota”, Sciencedirect.com https://www.sciencedirect.com/topics/immunology-and-microbiology/glomeromycota
Angela M. O’Callaghan. “Mycorrhizae”, Cooperative Extension https://www2.nau.edu/~gaud/bio300/mycorrhizae.htm
http://www.ersa.fvg.it/export/sites/ersa/aziende/monitoraggi/allegati/Patologia-I.pdf
https://www.koppert.it/sfide/controllo-delle-malattie/ticchiolatura-del-melo/
https://www.koppert.it/sfide/controllo-delle-malattie/botrite/
https://gd.eppo.int/taxon/SCLESC/photos
Peltier Angelique J., Bradley Carl A., Chilvers Martin I., Malvick Dean K., Mueller Daren S., Wise Kiersten A., Esker Paul D. 2012. “Biology, Yield loss and Control of Sclerotinia Stem Rot of Soybean”, Journal of Integrated Pest Management
Michailides T. J and Elmer P .A. G. 2000. "Botrytis gray mold of kiwifruit caused by Botrytis cinerea in the United States and New Zealand". Plant Disease
Paolo Gonthier, Ilenia Marafante, Giovanni Nicolotti, Jean-Claude Haudemand, Edi Pasquettaz, Giancarlo Zorzetto. “Difesa in foresta con urea contro Heterobasidium annosum”, regione.vda.it https://www.regione.vda.it/gestione/riviweb/templates/aspx/informatore.aspx?pkArt=160
“Heterobasidium annosum”, cabi.org https://www.cabi.org/isc/datasheet/27042
https://www.first-nature.com/fungi/heterobasidion-annosum.php
Stefania Cazzavillan. 2017. "Hericium erinaceus: un dono per il sistema nervoso", stefaniacazzavillan.it https://stefaniacazzavillan.it/hericium-erinaceus-un-dono-per-il-sistema-nervoso/
“Le numerose proprietà dell’Hericium”, micotherapy.it https://www.micotherapy.it/le-numerose-proprieta-dellhericium/
Arvind Bijalwan, KalpanaBahuguna, Amol Vasishth, Alankar Singh, Sumit Chaudhary, Anil Tyagi, MP Thakur, Tarun Kumar Thakur, Manmohan JR Dobriyal, Rajesh Kaushal, Arjun Singh, Nandini Maithani, Devendra Kumar, Gaurav Kothari, Pramod Kumar Chourasia. 2020. “Insights of medicinal mushroom (Ganoderma lucidum): prospects and potential in India”, Biodiversity International Journal
“Pleurotus ostreatus”, mondofunghi.it https://www.mondofunghi.com/pleurotus-ostreatus-funghi-pleurotus.html
Pleurotus ostratus – fungo gelone – ottimo commestibile”, funghimagazine.it https://funghimagazine.it/pleurotus-ostreatus-gelone-ottimo-commestibile/
S. M. Hosseini, K. Khosravi-darani and H. Nikoopour. 2009. Production of Mycoprotein by Fusarium venenatum Growth on Modified Vogel Medium, Asian Journal of Chemistry
https://puntodincontro.mx/2021/09/09/lhuitlacoche-la-gastronomia-del-mais-ammuffito/
https://www.vegolosi.it/news/quorn-il-fungo-principe-del-mercato-alimentare/
Ti è piaciuto l'articolo?
 BioDaily.it non riceve alcun contributo pubblico né ospita alcuna pubblicità, quindi si sostiene esclusivamente grazie alle donazioni dei lettori. Ti ringraziamo qualora tu volessi fare una donazione al nostro progetto, puoi farlo cliccando su questo messaggio.
BioDaily.it non riceve alcun contributo pubblico né ospita alcuna pubblicità, quindi si sostiene esclusivamente grazie alle donazioni dei lettori. Ti ringraziamo qualora tu volessi fare una donazione al nostro progetto, puoi farlo cliccando su questo messaggio.